




История происхождения и развития жизни
3.3.4.2. Эволюция в лабораторных условиях
Для выживания в кардинально меняющихся условиях окружающей среды популяции бактерий нуждаются в некотором запасе генетической изменчивости. То, что они обладают таковой, можно показать на примерах "лабораторной эволюции". Особенно легко могут быть получены невосприимчивые к антибиотикам мутанты в достаточно большой популяции на содержащих антибиотики питательных средах Наиболее ценно для дальнейших эволюционных опытов исследование катаболических изменений вещества (включающих распад субстанции) на основании простого опыта.
Устойчивость к антибиотикам
Благодаря флюктуационным тестам по Луриа и Дельбрюку, оказалось возможным косвенно доказать, что клетки, невосприимчивые к антибиотикам, существуют ещё до воздействия антибиотиком, и поэтому антибиотик едва ли пригоден как средство селекции. "Репликатеcт" по Ледербергу дает нам прямое доказательство этому.
Для понимания механизма возникновения сопротивляемости на молекулярном уровне нам необходимо привлечь в круг нашего внимания антибиотики, которые посредством присоединения к рибосомным протеинам тормозят синтез протеинов.
Устойчивость к антибиотику спектиномицину у E.coli взаимосвязана со структурой 5S белка малой рибосомной субъединицы. Мутация приводит к замене аминокислоты серия на пролин и, соответственно, к замене урацила на цитозин в информационной РНК Следствием этой замены является изменение вторичной структуры протеина, что затрагивает также и моста крепления спектиномицина. При этом антибиотик уже больше не может "нападать" на протеин 5S.
Еще одна возможность выработки устойчивости состоит, например, у бактерий штамма Escherichia coli в инактивации хлорамфеникола путем ацетилирования (присоединения остатка уксусной кислоты). Этому способствует деятельность фермента хлорамфеникол - ацетилтрансферазы (ХАТ). В этом случае ХАТ присутствует в клетке в очень малой концентрации. Приспособляемость к возрастающей концентрации антибиотика проявляется в увеличении числа резистентных генов (рис. 3.26), кодирующих ХАТ.
Таким образом, более высокие концентрации ХАТ могут быстрее инактивировать хлорамфеникол. Если уменьшается концентрация антибиотика, то число копий гена уменьшается. В описанных случаях формирования устойчивости к антибиотикам речь идёт о приспособлении к условиям окружающей среды, при этом организм не использовал предлагаемое вещество. Однако использование нового вещества открывает перед бактерией новое жизненное пространство. Об этом речь пойдет в следующем разделе.
Количество фермента часто контролируется геном-регулятором rw при помощи протеина (репреcсора) (рис. 3.27). Если имеется вещес-rBOW, подлежащее разложению, то, благодаря активности гена-регулятора гw, "запускается" феpмeнтный синтез, и ферменты, которые разлагают вещество w (А, В, С на рис. 3.27), образуются в больших количествах.
Если для расщепления необходима индукция фермента, то достаточно мутации по направлению к основному ферментному синтезу для усиления реакции замещения.
Плохо поддающееся ферменту вещество S можно лучше разложить, если в гене-регуляторе произойдет мутация гw --> гc. Ферменты А, В и С могут быть конститутивно, то есть постоянно, неконтролируемо синтезированы, даже тогда, когда вещество w отсутствует, хотя обычно оно необходимо для "включения" процесса расщепления. Например, количество фермента А (рис. 3.27) может в результате синтеза увеличиться тысячекратно. Это делает возможным освоение новых субстратов, для которых характерно небольшое различие молекулярного состава (рис. 3.28). Высокая концентрация фермента усиливает обычно слабое расщепление субстрата. Если же некоторое исходное вещество S, с трудом поддающееся разложению, превращается в продукт I, который, в свою очередь, может быть в дальнейшем использован в уже существующих процессах обмена веществ, то некоторое вещество, ранее почти не разлагавшееся, может даже стать единственным источником углерода (С). Описанную ситуацию как нельзя лучше подтверждает пример развития природного штамма Enterobacter aerogenes. Ксилит, как единственный источник углерода (С) недостаточен для его роста (рис. 3.29). Были найдены мутанты, которые использовали 0,2% ксилитную среду, то есть могли на этом расти. Эта способность была выработана благодаря дерепрессии рибитдегидрогеназы (РДГ) (то есть отмены репрессора инактивации). РДГ может приводить к реакции обмена ксилита, но не может быть им индуцирован, это означает, что ксилит не может вызвать синтез рибитдегидрогеназы. После мутации к констуитивному синтезу фермента ксилит, наконец, может быть превращен РДГ в Д-ксилулозу. Ксилулоза - это промежуточный продукт в Д-арабит-обмене веществ, в результате которого появляется новый источник углерода. А предпосылкой к этому служат две исходных реакции расщепления рибита и арабита.
Часто для нового расщепляемого вещества уже имеются готовые способы обмена. Готовые способы для его расщепления даются как бы "напрокат" и мутации для этого не требуются. Можно предположить, что именно это и происходит в случае расщепления гербицида хлоридазона Phenylobacterium immobile.
Ферменты с новыми активностями
Pseudomonas aeruginoza начала проявлять после мутации абсолютно новую субстратную специфичность. В этом случае можно наблюдать прямую лабораторную эволюцию амидазы (амидорасщепляющих ферментов). А-амидаза природного типа гидролизует (расщепляет) ацетамид и пропионамид:
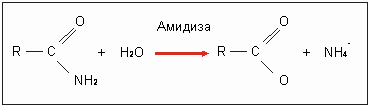 |
Продукты реакции могут использоваться как источники углерода(С) и азота (N). Оба вещества являются как хорошими субстратами, так и ферментными индукторами для амилазы. Это означает, что они легко расщепляются амидазами и вызывают усиленный синтез амидаз (рис. 3.30). Если они присоединяют амид, то происходит конформационное изменение, изменение объемной структуры амидаз, которое делает возможным катализ (расщепление). Бутирамид не может, однако, подвергаться гидролизу. Он подавляет в конкурентной борьбе имеющиеся индуцирующие амиды и даже синтез амидазы. Даже у констуитивных мутантов ферментный синтез подавляется (репрессируется) (ami R-). Если бутирамид присоединен к индукторным точкам фермента, то с изменениями в направлении активной конформации явно покончено.
Был изолирован мутант, являющийся новой амидазой, В-амидазой (рис. 3.30). Ею был гидролизован бутирамид, который мог служить источником С и N.
В результате дальнейших мутаций он превращается в валерамид. В-амидаза отличается от А-амидазы природного типа другой электрофорезной мобильностью. Положительная перекрестная реакция с антисывороткой очищенной А-амидазы показала однако, что оба фермента очень схожи друг с другом.
Расширение субстратной специфичности объясняется обменом серина на фенилаланин в позиции фермента (ami 46).
фенилаланин является более гидрофобным и объемистым, чем серии. Обмен аминокислот не вызывает изменений стабильности, разве что потерю потенциальной Н-связи, что, в свою очередь, привело к более высокой степени гибкости (упругости, эластичности) фермента, которая настолько изменила его конформацию, что и для бутирамида возникла возможность расщепления.
Впечатляюще была продемонстрирована "пластичность" жизни на примере EBG-фермента ("evolved b-galactosidase). Его можно рассматривать как идеальный пример "лабораторной эволюции". Возникновение,
новой функции можно проследить в данном случае в ходе постоянной селекции мутационно измененных структур фермента, являющегося непреложной частью нормально функционирующего организма.
Это становится возможным, если в клетку бактерии многократно заложены гены со схожими функциями, например, синтеза тирозина. В этом случае выживаемость популяции обеспечивается полным (тотальным) блокированием прежних привычных способов. Изначальная генетическая "оснащенность" бактерии создает потенциал для развития путем расщепления или синтеза новых соединений. На основе имеющихся способов обмена веществ
и имеющихся механизмов регуляции происходит посредством мутации и последующей селекции приспособление организмов (микроэволюция). Многообразие ферментов и их способность варьировать благодаря мутации дают возможность удалять органические соединения необычных структур (гербицидов, инсектицидов, нежелательных промышленных отходов). Такие ферменты особенно распространены в группе псевдомонад.
Мутанты с новыми свойствами служат также синтезу органических соединений в огромных количествах. Так глицерин-потребляющий мутант (Corinebacterium alkanolyticum) производит 72 г глутаминовой кислоты в парафиносодержащей среде в качестве единственного С-источника.
Заключение:
Генетический материал организмов обычно передается из поколения в поколение неизменным. В редких случаях происходят полностью неуправляемые, спонтанные мутации. Если условия окружающей среды резко меняются, то приспособляемость при помощи фенотипических модификаций становится невозможной, а определенные спонтанные мутации могут обеспечить выживаемость. В достаточно большой популяции всегда имеются наготове в различном количество мутанты для требуемой в данный момент адаптации. Возникновение подобных мутантов происходит в большей или меньшей степени задолго до того. Выработка сопротивляемости по отношению к антибиотикам, несмотря на столь обширные последствия, сводится к относительно небольшим изменениям (обмен аминокислот, потеря или ввод лишнего нуклеотида).
Для защиты от антибиотиков штамм бактерий пытается при помощи химических превращений дезактивировать их. Многие штаммы бактерий даже расщепляют антибиотики, как это происходит с антибиотиком хлорамфениколом, который может затем служить источником углерода и азота. Для повышения биохимической продуктивности зачастую бывает достаточно модификаций регуляции вследствие изменения репрессора или мест его связывания с геном-регулятором. Замена в аминокислоте фермента может настолько изменить структуру протеина, что это сузит его специфичность к субстрату. Первоначальный характер измененного фермента остается, тем не менее, ясно узнаваемым, что и показывает иммунология. Опыты в области лабораторной эволюции показывают, как может измениться исходная форма путем изменения генной продуктивности, в основании которой лежит замена нуклеотидов или увеличение участков генов.
Увеличение информации, полученное в результате мутации и селекции, существенно не нарушает границ жизненной формы. Новые свойства ограничены узкой сферой. Исходный тип распознаваем, и обратная мутация принципиально возможна.
Перечисленные преимущества, которые предоставляют для эволюционных опытов бактерии, подтверждают, однако, что все они остаются в области микроэволюции, подобно проводимому человеком на протяжении столетий разведению животных и растений, которое может быть сравнимо с образованием новых видов (сравните разделы 3.3.5; 3.4).
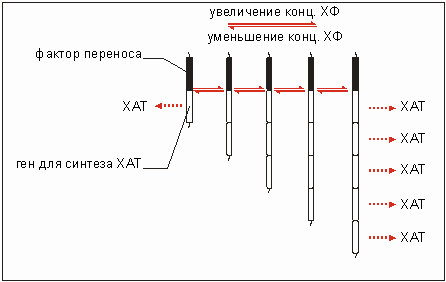
Рис. 3.26. Умножение гена в присутствии хлорам - феникола (ХМ) в возрастающей концентрации. ХАТ - хлорамфеникол - ацетилтрансфераза, фермент, дезактивирующий хлорамфеникол. При уменьшении концентрации последнего происходит уменьшение числа копий гена. Участок трансфер-фактора содержит гены, необходимые для переноса.
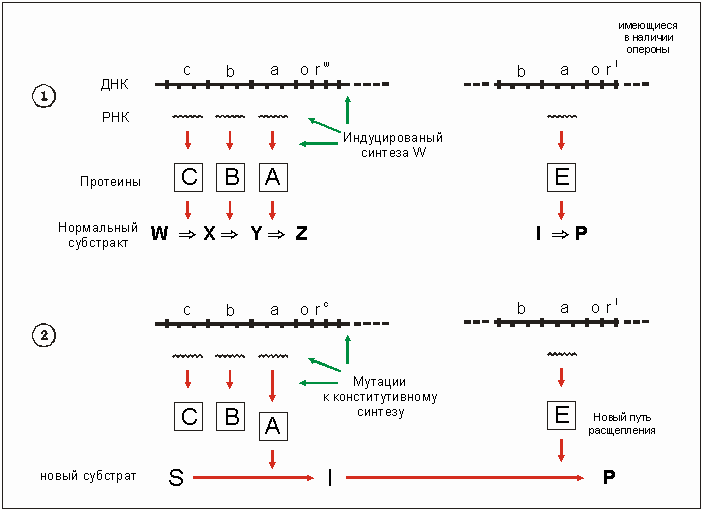
Рис. 3.27. Модификация уже имеющейся цепочки 1 - > Р с помощью заимствованного из другой цепи фермента. 1 - Первоначальная ситуация: две независимые друг от друга субстанции с сопутствующими реакциями обмена. 2 - После мутации к конститутивному синтезу синтезируется 6есконтрольно много фермента А (который обычно только превращает вещество У в Z). У него низкая субстратная специфичность по отношению к неиспользовавшемуся (незнакомому клетке) веществу S. Случайным образом оно превращается в вещество I, которое может в дальнейшем быть использовано в уже имеющейся цепочке обмена I - > Р. Благодаря крайне высокой концентрации фермента А, являющегося результатом мутации, формируется новый путь упрощения путей синтеза, который основан на совмещении существовавших прежде двух путей обмена веществ. (По MORTLOCK, 1982, в сокращении.)

Рис. 3.28. Изменение субстратного обмена посредством мутации к основной форме, фермент обладает небольшой субстратной специфичностью.
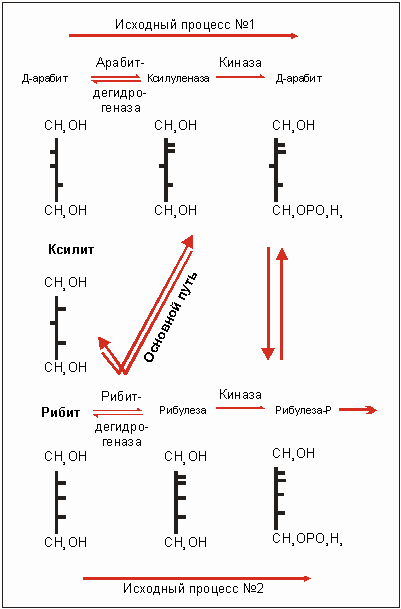
Рис. 3.29. Значение основного синтеза рибитдегидрогеназы для реализации ксилита. На основании двух исходных процессов обмена веществ и в ходе мутации к основной форме ксилит, непригодный к применению в обычных условиях, поддается обмену. Пояснения в тексте. (По Wu, Lin и Тanaka, 1968.)
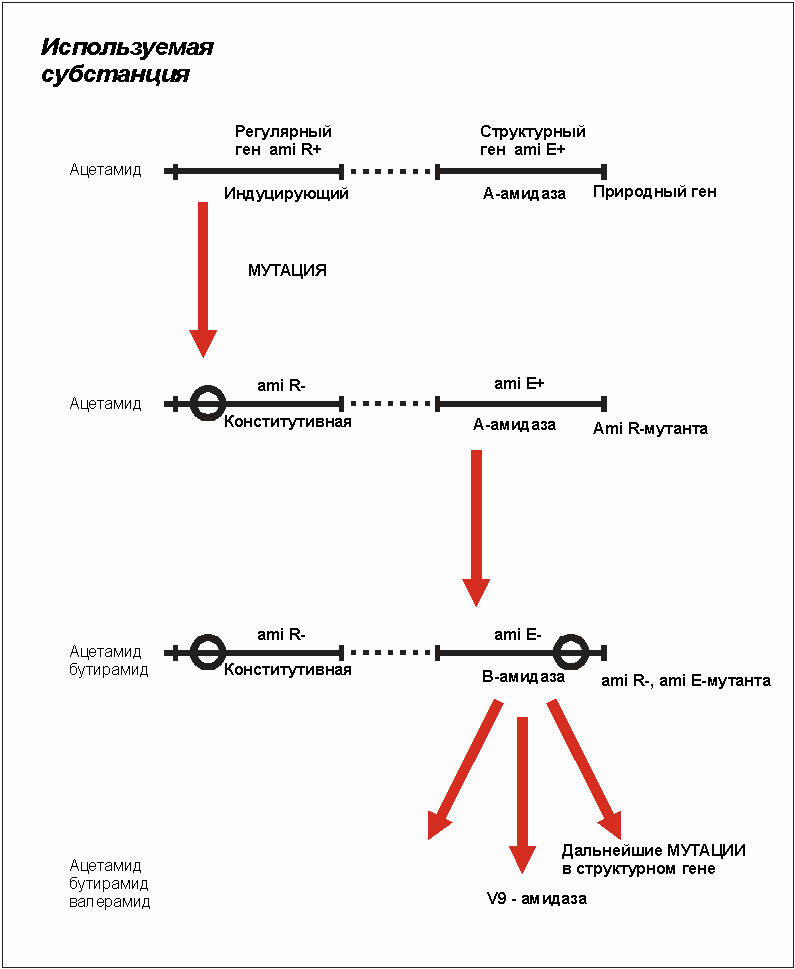
Рис. 3.30. Мутации для расширения использования
амидов. Пояснения в тексте. (По KAMPFE, 1981), с изменениями).



