




История происхождения и развития жизни
8.3. Геологические системы
Однотипность относительного расположения отложений пород по всей Земле.
Последовательность появления друг за другом ископаемых, связанных с отложениями того или иного периода, позволяет в основных чертах провести параллели между характером отложений по всей поверхности Земли. В порода* под слоями эокембрия находится так называемый докембрии (рис. 8.8). В них встречались до сих пор только бактерии, очень мало грибковых образований, организмов неизвестного происхождения, а также водоросли и многочисленные известняковые образования подобного характера. Отложения так называемого палеозоя, часто представляющие собой многокилометровые песчаники типа серой валки, песчаные и глинистые сланцы, а также известняки кембрийского, ордовикского и силурийского периодов и большей части девонского периода, содержат почти все без исключения морские организмы, такие как. например, трилобиты (см. рис. 8.3). Затем следуют породы карбонового периода с многочисленными превратившимися в уголь растительными остатками (см. раздел 8.5). Завершение отложений каменного угля приводит к значительным изменениям ископаемых других последующих отложений пород. Во многих случаях они принимают, выражаясь обобщенно, ватообразный или соответственно сходный с пустынным ландшафт (подобный встречался до этого только в девонских отложениях древнего красного песчаника в некоторых областях Северного полушария). Часто встречающиеся сухие илистые поверхности, обширно окрашенные в красный свет, представляют собой конгломератные песчано-мергелистые глинистые слои, которые можно опознать по часто встречающимся "законсервированным" следам (см. рис. 8.10), а также но появлению кристаллов соли. Отложения эпиконтинентального мелководного мори (моря, которое временами заливало части континента) пермского, триасового, юрского (раздел 8.6), а также мелового и третичного периодов залегают вперемежку с красными отложениями или выделяются из них. Эти морские известняки, мергелистые отложения, темные глины и песчаники погребают под собой богатую фауну. Принадлежность к тому или иному разделу этой системы определяется на основании находок в осадочных отложениях руководящей фауны, то есть ископаемых останков, характерных именно для этой части разреза. Требования к руководящим ископаемым следующие: частая встречаемость, легкая узнаваемость, краткое время существования вида, широкое территориальное распространение. Появление руководящей фауны в различных по составу и географически удаленных друг от друга отложениях пород (например, песчаниках и известняках) позволяет их коррелировать, то есть предполагается, что они образовались в одно и то же время в соответствии с законом о залегании пород. Таким образом, слои прород с одинаковыми руководящими окаменелостями представляют собой слои, относящиеся к определенным временным периодам, продолжительность которых по представлениям теории сотворения иная, чем по представлениям теории эволюции (см. разд. 8.6.5.).
Самое молодое и самое высокое горное образование было обнаружено в альпийских складках (см. рис. 8.11). Предположительно они были образованы сдвигом и столкновением литосферных плит. В рамках теории актуализма, которая пытается проецировать процессы современности на прошлое и, основываясь на наблюдаемых в настоящее время тектонических движениях плит, создать модели исторического прошлого Земли, попытки подобного толкования носят чисто умозрительный характер. В настоящее время можно проследить за медленным движением континентальных плит. Однако невозможно доказать, допустимо ли в данном случае говорить о более или менее постоянном движении, которое одинаково равномерно происходит на протяжении сотен миллионов лет, или же речь идет о фазах затухания более значительных в прошлом движений земной коры. Наряду с доказательствами быстрого вертикального движения земной коры (например, образование бассейнов каменного угля - см. раздел 8.5.4) в истории развития Земли имеются примеры и существенно более быстрого горизонтального движения. Как было уже сказано в главе 1, обрабатывающиеся в настоящее время данные могут иметь абсолютно разное толкование.
Последовательность отложения пород в ходе исторического развития Земли изображена в стратиграфической таблице (см. табл. 8.1). Если принимать во внимание широкомасштабное применение закона о залегании пород (распространение его на любую точку поверхности Земного шара, в сочетании с фактом наличия руководящих окаменелостей), то можно считать эту таблицу достаточно достоверной.
Последовательность отложения ископаемых
Известная закономерность отложения пород в рамках эволюционной теории и теории сотворения интерпретируется абсолютно по-разному.
Толкование в рамках эволюционной модели. Ископаемые остатки, идентичные по всему разрезу осадочных отложений, свидетельствуют о длительном всеобщем процессе развития органического мира. Отсутствие ископаемых остатков животных и растений, известных в настоящее время, например, в породах палеозойского и мезозойского периодов объясняется тем, что эти виды тогда еще просто не существовали. Например, останки плацентарных млекопитающих можно обнаружить исключительно в третичном периоде, т. к. они должны были развиться лишь к этому времени. Все сказанное выше подходит и для других групп организмов.
Трудности толкования в рамках эволюционной модели. Основной проблемой является необходимость признания факта медленного образования слоев земной коры, содержащих ископаемые. Без этого эволюционное толкование невозможно. Об ископаемых, которые, но мнению авторов, свидетельствуют об их образовании в результате катастрофы, смотрите в разделах 8.5 (каменный уголь) и 8.6 (верхнеюрские известняковые плато). Кроме того, при эволюционном толковании предполагается наличие огромного числа переходных форм между основными типами организмов. В разд. 8.7 подробно говорится о том, что таковые до настоящего времени не обнаружены. Иногда некоторые ископаемые находки пытаются представить в качестве промежуточных звеньев, однако скоро становится ясно, что подобное толкование можно оспорить. Отсутствие промежуточных звеньев можно объяснить лишь тем. что все эволюционные процессы протекали в настолько малых пограничных популяциях, что вероятность их превращения в ископаемые слишком мала (см. раздел 8.7.9).
Толкование в рамках модели сотворения. После того как Земля была опустошена в результате всемирного потопа (см. раздел 8.5.5.) новое заселение могло происходить неоднократно. Распространившимся по всему свету новообразованным сообществам живых существ (биоценозам) были свойственны высокая плодовитость и непродолжительное время жизни. Можно предположить, что они по прошествии небольшого периода времени распадались и заменялись, другими. Известно, что современные сильно измененные экологические районы имеют тенденцию к изменению в течение короткого периода времени. Сначала происходит смена одних кратковременно существующих сообществ другими (сукцессия) до тех пор, пока не достигается конечная стадия (климакс) с ее разнообразием биоценоза.
Примером подобного заселения живыми существами могут служить фазы восстановления лесного массива после вырубки: сначала восстанавливается поросль кипрея, за ней следует сообщество песчаного вейника, оно сменяется зарослями осины, ивняка и березняка, которые, в конце концов освобождают место для дубовых и буковых лесой (по SCAMONI, 1963, с, 137). Факторами, обуславливающими сукцессию, являются: структура почвы, различная всхожесть ни местах, незанятых растительностью, возможности размножении, условия конкуренции с соседями и т.д.
По такой же схеме могло происходить отложение ископаемых морского мелководья. Швен (1988) предполагает возможность, более или менее синхронно протекающих (мегасукцессионных) процессов смены биоценозов в масштабе планеты. После гибели всей земной биосферы в результате глобальной катастрофы именно эти процессы и получили развитие, завершившись формированием сообществ организмов третичного и четвертичного периода. Поскольку многие отложения мелководных частей моря содержат известь, то часто в них содержатся микроскопические частички ископаемых, которые указывают на значительное преобладание массы отдельных морских организмов, для развития которых имелись благоприятные условия. Подобный ход развития мог привести к значительному увеличению числа моллюсков, рыб и т. д. Поскольку во всем мире исходные условия для сукцессии были одинаковыми, то следствием этого явилось изначально схожее в мировом масштабе явление захоронения живых сообществ, которые снова и снова "консервировались" в ходе процессов захоронения, носивших характер катастроф (см. раздел 8.6.5) и были последовательно друг за другом погребены. Одинаковое появление и исчезновение различных групп организмов в "летописи" ископаемых объясняется большей частью экологическими причинами, что в известной степени дает возможность проверить эти гипотезы.
Широко распространено предположение, что следующие друг за другом во времени биоценозы со своими расположенными в произвольном порядке системами организмов находят отражение в чередовании окаменелостей, образовавшихся в ходе исторического развития Земли (см. табл. 8.1). Их возникновение могло быть обусловлено, кроме всего прочего, следующими факторами:
- наличном в период после всемирного потопа жизненного пространства (сначала это были, вероятно, только морские, затем пресноводные, и только потом во все возрастающем объеме материковые ареалы обитания),
- временной период смены генераций и максимальная скорость распространения,
- разнообразные экологические причины,
- многочисленные, однако с течением времени затухающие катастрофы геологического и биологического характера, которые привели к мгновенному переходу в ископаемые, к консервации их и к своего рода "моментальной фотографии" тех или иных ареалов обитания,
- распространение выживших в результате катастроф популяций и распространение организмов в эти подвергшиеся воздействиям катастроф области.
Трудности, встречающиеся при попытках толкования этих гипотез в рамках модели сотворения Наиболее уязвимое место мололи сотворения в том, что за основные типы живых существ должны были быть созданы одновременно. И хотя на основании мега-сукцессионной модели невозможно по экологическим причинам ожидать в породах отложений, например, мезозойской эры присутствия окаменелостей современных форм, ясно, что одновременно г ареалом обитания древних рептилий должен был существовать биотоп с привычными для нас формами. То есть среди ископаемых мезозойской эры, пусть даже в качестве редкой находки, должны встречаться останки этих форм и даже человека.
Сообщения о подобных находках публикуются постоянно. В качестве примеров можно перечислить следующие факты: одновременное появление следов динозавров и человекообразных в меловом периоде, находки человеческих останков в отложениях третичного периода (миоцен), обнаружение цветочной пыльцы высокоразвитых растений в докембрийских отложениях (рис. 8.12). Следует упомянуть также о частых случаях залегания слоев в обратном порядке, когда более "древние" отложения оказываются над более "молодыми", если судить об их возрасте по найденным в них органическим останкам. В ряде случаев такое необычное залегание пород никак нельзя объяснить тектоническими сдвигами. В постановке проблемы проявляется известное сходство. Подобно тому, как подвергаются сомнениям промежуточные звенья в эволюционной модели, сообщения о явлениях этих нетипичных по очередности расположения слоев пород подвергаются сильным нападкам отчасти и со стороны приверженцев учения о сотворении. В таких случаях необходима тщательная проверка учеными, придерживающимися различных точек зрения. Но даже если некоторые из этих сообщений будут признаны достоверными, ясно, что подавляющее большинство палеонтологических данных не содержит этого нетипичного расположения останков фауны и флоры. Обычно преобладает нормальная последовательность слоев отложения. Это и требует объяснения в рамках теории сотворения. Следует предположить, что существа, вначале типичные лишь для третичного периода, пережили и выдержали катастрофические перемены в виде очень небольших групп Вероятно, они могли сильно размножиться и распространиться в мировом масштабе лишь в более поздний и благоприятный для них период мегасукцессии (третичный). Именно тогда, видимо, и мог происходить процесс их перехода в отложения. Вероятность минерализации для представителей небольших популяций настолько мала, что их останки не сохранились. Такие малые сообщества имеют очевидное сходство с постулируемыми в эволюционной теории сверхмалыми и также не попавшими в число окаменелостей переходными пограничными популяциями, в которых должна протекать так называемая "быстрая эволюция" (образование новых крупных таксонов). Об этом см. в разд. 3.5.3 (пунктуализм). Доля иронии заключается в том, что для защиты основных спорных вопросов как в теории сотворения, так и в эволюционной теории используется одна и та же аргументация.
Признание факта всемирного потопа как постулат, принимаемый на веру
Итак, модель единовременного создания живых организмов, процесс дальнейшего распространения и выживания которых контролировался экологическими катастрофами, противопоставляется модели медленного, длительного по времени процесса эволюции. Решение, какая из них справедлива, зависит от того, сколько времени требовалось на образование осадочных отложений.
Структура отложений пород дает явные доказательства катастрофического характера их образования. Но на этом уровне невозможно сделать вывод о причинах катастроф. Если в этой книге предположить, что причиной всего этого был библейский всемирный потоп, последствия которого сказывались еще многие столетия спустя во все менее значительных "затухающих" катастрофах, то авторы вполне сознательно подводят читателя к этой проблеме, чтобы дать ему в большей степени почувствовать "аргументационный водораздел" (см. раздел 8.5.5).
Модель сотворения, опираясь на данные радиоактивного распада и ряд других доказательств, предполагает образование отложений в короткий промежуток времени. Поэтому, вероятно, имеет смысл обсудить здесь методики определения возраста, основанные на радиоактивном распаде.

Рис. 8.6. Слои осадочных пород в районе Цюриха.

Рис. 8.7. Слои осадочных пород в районе Цюриха. Те же слои, что и на рис. 8.6, но в более крупном масштабе. (Scheven, 1986, с. 23.)

Рис. 8.8. Происходящая в процессе отложения деформация слоек (докембрий, Апелькрос, Шотландия). Деформации слоев возникают непосредственно после того, как произошел процесс, отложения {зачастую во время дальнейшего оседания отложений), в состоянии, еще подверженном изменениям. Причиной подобных деформаций могут служит!, напряжения, вызванные различными нагрузками или разрушения, вызванные внезапными толчками и земной коре. Существенное условие, необходимое для образовании подобных структур, - предшествовавшее этому быстрое отложение пород. Отпечатавшиеся докембрийские слои. Следующие по порядку один за другим (ширина образования 70 см), являются лишь малой частью огромных, более сотни метров, слоев, почти сплоим, деформированных, которые занимают пло-щад1, многих квадратных километров.
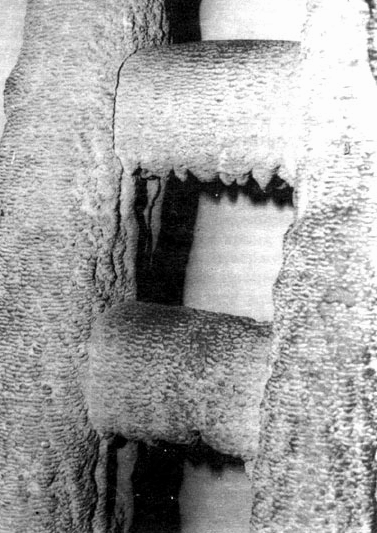
Рис. 8.9. Быстрое образование известняковой конкреции на лестнице в горячем водоеме ок. 5 см толщины. (Музей Бергау, Бохум.)

Рис. 8.10. Поверхность пласта с сетью окаменевших следов (отпечатки в негативном изображении) и следы лап ископаемых рептилии Chirotherium н пестрых песчаниках (нижний триасовый период). Обрывки сетчатых отпечатков возникают к том случае, если высохшие отложения растрескиваются (как глинистые лужи при высыханий) и щели ВНОВ1, покрываются и заполняются свежими отложениями. Так как высохшие расщелины и отпечатки выветриваются, то сохранение подобных структур в качестве ископаемых доказывает достаточно быстрое наслоение новых отложении. При этом возникает новый оттиск - негативный. (Палеонтологический музей университета, Тюбиген.)

Рис. 8.11. Складчатые отложения в Альпах. (Бриенц, Швейцария.)
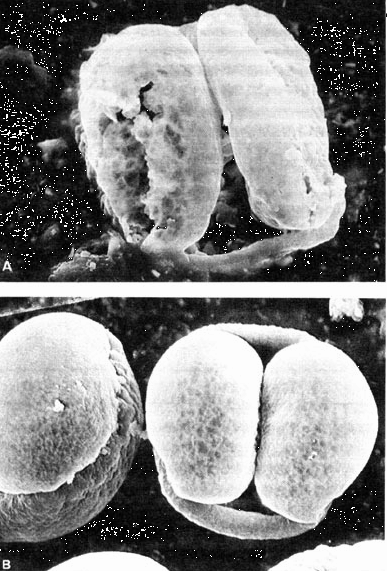
Puc. 8.12. Изображения пыльцы из докембрийских пород (А) и пыльцы современных растений (В), полученные на электронном микроскопе,. Появились разного рода сообщения о том, что отложения докембрийского периода содержат следы пыльцы и даже останки цветковых растении. На этих снимках изображены: А - пробы пыльцы, взятой из докембрийской формации, В - современная пыльца. Разные пробы брали из соответствующего горизонта раскопок, и только в одном-единственном случае в пробе не было обнаружено пыльцы. Были предприняты самые строгие меры предосторожности, призванные предотвратить загрязнение проб современной пыльцой, по все повторяющиеся доказательства наличия пыльцы сосны не позволяют объяснить этот факт простым загрязнением. (По Е. WILLIAMS и др., Creation Research Society Quarterly, 24(4), March, 173-182.)



